检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
 下载Firefox
下载Firefox
检测到您当前使用浏览器版本过于老旧,会导致无法正常浏览网站;请您使用电脑里的其他浏览器如:360、QQ、搜狗浏览器的极速模式浏览,或者使用谷歌、火狐等浏览器。
下载Firefox
2022年9月15日,js33333金沙线路检测季雄课题组在Molecular Cell杂志在线发表了题为“Targeted protein degradation reveals RNA Pol II heterogeneity and functional diversity”的研究论文。该研究系统阐释了真核生物细胞中RNA聚合酶II各亚基对转录和转录后过程的直接调控作用和潜在机制,首次表明RNA聚合酶II是一个优化酶(Optimized polymerase, OPPO), 其亚基在转录和转录后过程中发挥着不同,甚至特异的作用。

RNA聚合酶II主要负责蛋白编码基因的转录,是由12个亚基组成的蛋白复合体。领域内一直认为RNA聚合酶是作为一个整体发挥功能的全酶(holoenzyme),由于组成RNA聚合酶的亚基是必需基因,传统干扰手段处理时间较长,不可避免地会引入二级效应,因此其亚基的直接调控功能不清楚而且被长期忽视。此外令人困惑的是,RNA聚合酶亚基应该是组成性表达的,但是其失调和变异常常会伴随不同组织中的不同疾病,特别是脑部疾病。
研究者首先在小鼠胚胎干细胞中建立了RNA聚合酶II 12个亚基的瞬时降解细胞系,联合染色质基因组与蛋白质组分别定量各个亚基的染色质互作基因组位点和蛋白质。结果显示各亚基在基因组范围内呈现非均一占位、互作蛋白存在显著差异,例如RPB4与RPB7相较于其他亚基,其与转录延伸调控蛋白NELF复合物的互作强度明显较低。结合定量质谱和分子筛生化分析,研究者同样发现了各亚基在转录过程中呈现非均一的互作甚至可能存在从全酶解离的过程。
接着研究者使用新生RNA测序实验(PRO-Seq)探究各亚基瞬时降解后(1h)对新生RNA生成的影响。有趣的是不同亚基的降解可产生不同程度的转录失衡,而一些亚基如RPB4、RPB9、RPB11、RPB12的降解不影响新生RNA生成,表明这些亚基瞬时降解后,不影响转录过程。与此同时,研究者继续使用染色质RNA测序实验(ChAR-Seq)对部分亚基降解后转录失衡现象进行了验证,得出相同的结论,表明数据的可靠性。随后,研究者通过WGCNA分析,将RNA聚合酶II复合物各亚基在染色质定位情况与降解后不同基因的转录情况进行了关联性分析,发现亚基在基因的结合强度越高,该基因的转录越依赖该亚基,暗示亚基对不同基因转录调控存在基因特异性。
为了进一步证实上述猜想,研究者开展了长时程降解(12h)后的PolyA-RNA-Seq实验,探究各亚基对不同基因成熟mRNA的影响。研究者发现RPB1、RPB2、RPB5、RPB6、RPB7、RPB8的降解可导致大部分基因转录下降,表明这些亚基参与RNA聚合酶全酶稳定性,这与免疫荧光成像实验中这些亚基降解后导致核内全酶显著降低的结果相呼应,然而RPB4和RPB12的降解在多个独立实验中均没有呈现变化,研究者认为它们可能存在蛋白剪接变体代偿了亚基降解的影响。有意思的是RPB3、RPB9、RPB10和RPB11的降解仅造成部分基因转录下降,免疫荧光成像实验显示延长降解时间,核内仍然保留部分RNA聚合酶II复合物,暗示这类亚基在全酶中起调控作用。
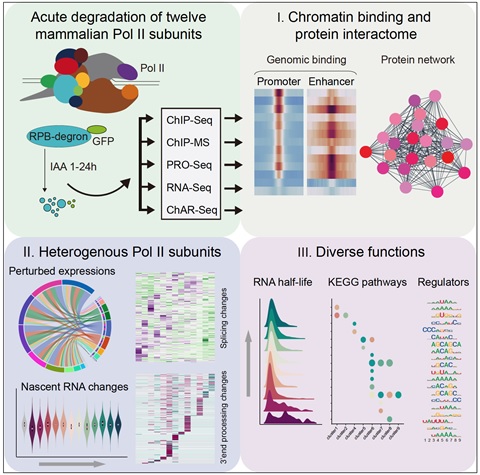
图1. 多学科实验技术和生物信息学分析,系统阐释了真核生物细胞中RNA聚合酶II各亚基对转录和转录后过程的直接调控作用。
PolyA-RNA-Seq实验结果同时显示一些亚基降解后可导致部分基因不同程度的表达上调,研究者分析基因上调可能是转录后过程的失调造成的,如转录本3’末端加工缺陷,剪接加工缺陷等。使用前人开发的Dogfinder软件,研究者鉴定出亚基降解前后存在转录通读的基因类别,随后结合基因3’末端加工系数的计算,发现RPB1、RPB3、RPB5、RPB8降解后可显著增加转录通读,这与Robert Roeder实验室2020年在Molecular Cell发表文章发现RPB1-CTD降解后造成基因通读的结果相吻合。同时研究者通过rMATs对转录剪接异常的基因进行鉴定,同样发现不同亚基的降解可造成不同基因的剪接异常。更深入地,对造成转录、转录后3’末端加工和剪接异常的潜在因素进行探索后,发现如DNA序列、RNA结合蛋白和RNA二级结构等多种因素参与决定亚基特异性的调控。

图2. 中国剪纸十二生肖代表RNA聚合酶II的12个亚基,占据在全酶复合体结构中的相对位置,构成了最吉祥的汉字“福”。 该研究报道了12个亚基优化RNA聚合酶II功能的基础。
RNA聚合酶亚基工作中,js33333金沙线路检测季雄研究员是该论文的通讯作者,js33333金沙线路检测李圆君博士和黄捷博士是该论文的共同第一作者。js33333金沙线路检测博士研究生朱峻毅、包丽君,西湖大学js33333金沙线路检测郭天南、朱怡课题组等为该工作提供了重要帮助。
2022年9月16日js33333金沙线路检测季雄课题组联合定量生物学中心齐志课题组在iScience杂志上发表了题为“CTCF DNA binding domain undergoes dynamic and selective protein–protein interactions”的研究论文,该文章揭示了CTCF的DNA结合结构域(DNA-binding Domain,DBD)能以不依赖无序结构域的方式形成动态的自聚集簇,在三维细胞核中能和绝缘子相关蛋白共聚集、和活跃转录因子不共定位,这一发现为解析CTCF的绝缘机制提供了一种全新的角度。

CTCF(CCCTC-binding factor)是脊椎动物体内目前已知的最主要的绝缘子结合蛋白,其可以阻断增强子对启动子的激活,抑制基因的表达,也可以作为“屏障”阻止异染色质的扩散。染色质构象捕获技术结果显示CTCF能促进染色质环结构的形成, 通过与Cohesin形成染色质环,折叠成有助于增强子和启动子互作的三维染色质构象。但在三维细胞核中CTCF如何阻碍染色质环内的增强子对环外基因启动子的激活,以及CTCF的各个结构域和绝缘子功能有怎样的关系鲜有报道。
研究者利用光诱导蛋白聚集的“optoDroplet”系统筛选到CTCF的DNA结合结构域能发生聚集;并在体外验证CTCF的DBD能聚集形成动态的液滴;接下来研究者通过蛋白共定位分析结合活细胞追踪成像,发现CTCF DBD的聚集能招募绝缘相关蛋白CHD8和BRD2,同时排开转录激活因子OCT4;通过对已发表的CTCF ChIP-Seq数据进行分析,研究者发现基因组上CTCF的基序越多,绝缘能力越强,并在表征绝缘活性的荧光素酶报告系统中插入不同数目的CTCF结合基序进行了验证。锌指结构是经典的结合核酸的蛋白结构域,为进一步探索核酸对CTCF DBD聚集的影响,研究者通过在体外相变体系加入核酸,在细胞实验上利用破坏核酸结合能力的突变体等证明了核酸抑制CTCF DBD的聚集。此外,生物信息学分析发现CTCF DNA结合结构域的精氨酸存在大量突变,该突变会影响CTCF DBD的DNA结合和绝缘能力。总之,研究者首次发现CTCF的DNA结合结构域可以发生自聚集,后续也验证了其它转录因子例如BCL6、GATA3、YY1的DNA结合结构域也能形成聚集,并能和功能相关蛋白共定位,同时P53蛋白的DNA结合结构域内癌症相关突变体R175H能降低其聚集体的稳定性,这些结果表明DNA结合结构域的聚集可能是一种较为普遍的转录因子的功能调控机制。
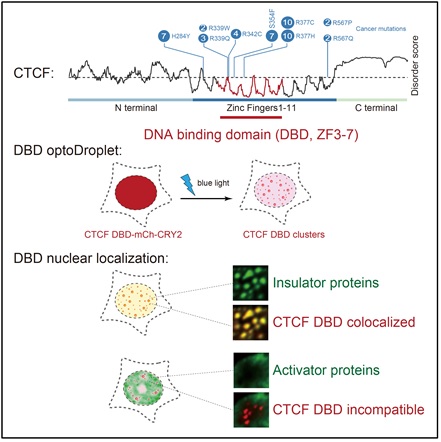
图3. CTCF的DNA结合结构域能和绝缘相关蛋白发生动态和特异性的相互作用
CTCF绝缘机制工作中,js33333金沙线路检测季雄研究员、定量生物学中心齐志研究员为本论文共同通讯作者。js33333金沙线路检测周蓉博士、田凯博士、黄捷博士为本论文共同第一作者。js33333金沙线路检测博士研究生段文嘉、付红烨,上海科技大学生命科学与技术学院马涵慧教授和冯颖等为该工作提供了重要帮助。这两个工作得到科技部国家重点研发计划、启东创新基金、国家自然科学基金、北大-清华生命科学联合中心和细胞增殖与分化教育部重点实验室的资助。感谢js33333金沙线路检测凤凰工程多个仪器平台对本项目的大力支持。
季雄课题组长期从事RNA聚合酶亚基功能调控和疾病机理的研究。主要集中在RNA聚合酶相关的新型功能、脑部疾病和核酸适配体筛选等方向,近期成果发表在Molecular Cell、 Nature Communications、Genome Biology、Cell Discovery、CMLS和iScience等杂志上,为选择性基因表达调控提供新的假说。现因发展需要,诚邀博士后和研究生加入。
原文链接:
1. https://www.cell.com/molecular-cell/fulltext/S1097-2765(22)00812-7
2. https://www.sciencedirect.com/science/article/pii/S2589004222012834
地址:北京市海淀区颐和园路5号
金光生命科学大楼
电话:010-62757794

北大生科官方微信

生声不息公众号